Tämä on vain kertauksena reologisen puolen kaskadeista, koska MMP-TIMP-kaskadi kylkii siihen.
- Reologisia yksityiskohtia: Rheological detail (Factor Va, FVia, activated C-protein)
REOLOGIAN alalta löysin uuden väitöskirjan lokakuun viime päiviltä.24.10.2009
Kjellberg Ulla. Factor V Leiden mutation and pregnancy. Haemostasis during pregnancy in non-carriers and carriers of factor V Leiden mutation, with special emphasis on placenta-mediated and venous thromboembolic complication and on blood coagulation and fibrinolysis markers for prediction of complication.ISBN 978-91-628- 7904-4 ( 30.10.2009).
- Abtraktissa, tiivistelmässä, mainitaan seuraavaa:
Faktori V Leiden-mutaatio nostaa tromboembolisten laskimoperäisten tukosten riskiä (VTE, venous, thromboembolism) yleensäkin. Tiedetään myös, että raskauden aikana hemostaattinen tasapaino siirtyy normaalistikin hyperkoagulabiliteetin suuntaan, mistä seuraa kohonnut VTE-insidenssi eli laskimotukosten kohonnutta esiintymistä Tässä väitöstyössä seuratiin 42 naisen raskauden aikaa (pregnancy) ja lapsivuodeaikaa (puerperium) .
APC (proteiini joka aktivoituu trombiinista ja trombomoduliinista)
Klassisen ja modifioidun
aktivoidun-C proteiinin (APC) suhteet laskivat raskauden aikana.
Kuitenkin modifioidun suhde nousi normaaligeenisille tyypillisen alarajan yläpuolelle ja sitä voidaan käyttää havaitsemaan FVL-mutaatio raskauden aikana.
Protrombo
Kohonneet pitoisuudet fragmentteja 1+2
( F1+2), liukoista fibriiniä (
soluble fibrin SF) ja
D-dimeerejä viittaavat veren koagulaation aktivoitumiseen.
Lisäksi nousee seuraavat tekijät:
fibrinogeeni,
faktori VIII,
PAI-1 ja
PAI-2 ( plasminogeenin aktivaattorin inhibiittorit 1 ja 2) .
Seuraavien tekijöitten pitoisuudet laskivat: vapaa
proteiini S,
t-PA aktiivisuus ( kudosperäinen plasminogeenin aktivaattori, tissue plasminogen activator activity).
Varsinaisen
Proteiini C:n -pitoisuudet säilyivät muuttumattomina.
Sonoclot-analyysi viittasi
raskauden aikaiseen hyperkoagulaatiotaipumuksen. Sonoclot-käyrän samoja viitearvoja voidaan käyttää koko raskauden aikana.
Tutkimuksessa tyypattiin
FVL-geeni 5986 naiselta. FVL kantajuuden prevalenssi oli 8.1%.
500 FVL geeninmutaation kantajaa ja 1058 kontrollia seurattiin pitempään ja hemostaassin merkitsijöitä rekisteröitiin. Mitään eroja ei havaittu istukkavälitteisissä komplikaatioissa raskauden kestoajassa. Seuraavien komplikaatioitten esiintymä oli matala: neonataali asfyxia, eklampsia, kohdunsisäinen sikiökuolema , synnytyksen aikainen sikiökuolema tai hyvin myöhäisen vaiheen selittämätön abortoituminen.
Yli 1000 ml verenmenetys synnytyksen aikana oli harvinaisempaa FVL-kantajuudessa.
FVL-kantajilla oli 3 tapauksessa syvä laskimotulppa (VTE) ja muilla ei ollut yhdessäkään tapauksessa. Pinnallisissa tromboflebiiteissä ei ollut ryhmien välillä eroa. On kyseenalaista genotyypittää FVL:n suhteen niitä terveitä naisia, joilla ei ole ollut laskimotulppia eikä genotyyppaus ole oikeutettua obstetrisen komplikaationkaan perusteella.
Raskauden aikana nousee
F1+2 ja
D-dimeeri ja pitoisuudet ovat korkeampia, mitä kahdeksan viikkoa synnytyksen jälkeen.
Liukoisen fibriinin
(SF) muuntelut olivat vähäisiä tai niitä ei esiintynyt. Sekä FVL-kantajuudessa että kontrolleilla F1+2 ja SF olivat samoilla tasoilla raskauden aikana.
FVL-kantajuudessa oli
D-dimeeripitoisuudet korkeammat sekä raskauden että lapsivuoteen aikana. Kaikkien merkitsijöiden pitoisuudet olivat samoilla alueilla naisilla, joilla oli tai joilla ei ollut komplikaatioita, eikä niihin vaikuttanut zygoottisuus tai lisänä ilmenevä trombofilia.
F1+2, SF tai D-dimeeri eivät siis pysty toimimaan FVL-kantajuudessa mitenkään plasentaperäisten komplikaatioitten tai syvän laskimotulpan riskin ennusteena.
- Väitöskirjan tausta-materiaalissa
selvitettiin kuvan avulla koagulaation ja antikoagulaation sekä fibrinolyysin asemat toisiinsa. Verrattuna aiempiin paradigmaattisiin kaavakuviin tekijä oli selventänyt muutamia yksityiskohtia.
PLASMA sisältää korkean pitoisuuden
proteaasi-inhibiittoreita. Sen takia useimpien
seriiniproteaasien (koagulaatiotekijöitten) aktiivi muoto pystyy vaikuttamaan vain hyvin pienen aikaa, koska puoliintumisaika on äärimmäisen lyhyt, sekuntteja. Poikkeuksena on seitsemäs tekijä
fVIIa, jota on aina hivenmääriä aktiivimuodossaan kiertämässä.
NORMAALI ENDOTEELI, verisuonten sisäpinta, pitää yllä veren juoksevuutta, fluiditeettiä estämällä (inhiboimalla) veren koaguloitumista ja
verihiutaleitten eli trombosyyttien aggrekoitumista, kasautumista röykkiöön.
VERISUONEN VAURIOITUMINEN on tila, jossa veri altistuu endoteeliä syvemmällä kudoksessa oleville tekijöille
, kudostekijälle , adhesiivisille ( takertuville) proteiineille kuten kollageenille, fibronektiinille, laminiinille, vitronektiinille ja
vW-faktorille ja ne alkavat edistää verihiutaleitten eli trombosyyttien adheesiota, liimautumista vauriokohtaan, ja alkaa koagulaatio.
PRIMÄÄRI HEMOSTAASSI, käsittää suonen supistumisen, trombosyyttien aktivoitumisen:
gpIb liittyy
vW-faktoriin endoteelisoluissa.
gpIIb tekee siltaa
gpIIIa:han.
Sitten
IIbIIIa vaikuttaa että trombosyytit pystyvät liittymään toisiinsa fibrinogeenillä ja muodostuu alkutulppa,
primary platelet plug.
SEKUNDÄÄRINEN HEMOSTAASSI varmistaa paremman vuodon tukkeamisen. Tässä osallistuvat proteolyyttiset entsyymit. Nämä koagulaatioentsyymit kiertävät normaalissa verenkierrossa aktivoitumattomina (
inertteinä) . Ne aktivoituvat proteolyyttisellä pilkkoutumisella (kuin ketjureaktiona) sitouduttuaan spesifiseen proteiinikofaktoriin fosfolipidipinnalla.
Siis näissä koagulaatiotekijöissä oli yksi, joka ei ole kiertävässä veressäkään täysin inertti, vaan pysyttelee hivenmäärissä aktiivisena (
faktori VIIa, joka tekee kompleksin
kudostekijän TF kanssa).
KUDOSTEKIJÄ
TF, tissue factor on kalvoproteiini, jota on adventitian fibroblasteissa.
KOAGULAATIOTEKIJÖISTÄ
Kun verisuonen pinta vaurioituu, TF pääsee sitoutumaan ohikulkevan veren faktoriin VIIa. Nyt muodostuva
kompleksi TF /;VIIa alkaa aktivoida erästä tekijää, joka sanotaan kymmenenneksi faktoriksi
F X. Se aktivoituu
F Xa muotoon.
Tämän tekijän kofaktori on
F V, joka aktivoiduttuaan on
F Va.
Nämä kaksi yhdessä (FXa ja FVa) alkavat aktivoida
protrombiinista (FII) esiin
trombiinia ( FIIa) .
Tätä alkua sanotaan ”
TF pathway”- tieksi , kudosfaktori-tieksi , ja se on koagulaation aloittava jakso (
initial phase).Mainittu kompleksi pystyy aktivoimaan myös
F IX-tekijän( yhdeksikön)
TROMBIINI sitoutuu verihiutaleitten,TROMBOSYYTTIEN , fosfolipidipintaan, jos siinä on Z-proteiinia. Tästä seuraa oikein
”thrombin burst”, amplifikaatio, kiihtyvä trombiinin muodostus. Trombiini pystyy aktivoimaan
tekijät F V, F VIII, F XI, ja F XIII (“kolmetoista”) ja samoin trombosyytit ja stimuloi trombosyyttejä erittämään vaikuttavasisältöisiä jyväsiään.
FVIII a ( aktiivi kahdeksikko) aktivoi
FXI (yhdentoista)
ja se aktivoi
FIX (yhdeksikön).
FVIIIa toimii kofaktorina sitten
FIXa kanssa. Tämä on ”
a tenase complex”, joka taas aktivoi tekijän FX.
Tällainen sykli jatkuu ja vaikuttaa, että yhä enemmän TROMBIINIA muodostuu.
TROMBIININ primääri rooli on muuttaa
liukoinen fibrinogeeni fibriiniksi. ( Mutta katsoa myös että fibrinolyysille tulee jonkinkaltaista säätelyä ,.estoakin: Trombiini aktivoi TAFI:n tässä vaiheessa. Trombomoduliinn läsnäolosta sitten tulee lisää aktivaatiota TAFI:lle. Kun on plasmiinin valmistumisvaihe plasmiini edelleen aktivoi TAFI - tekijää, fibrinolyysin inhibiittoria. TAFI ei omaa fysiologista inhibiitoria)
FIBRIINI on hemostaattisen tulpan pääkomponentti.
Fibriininmuodostuksen viimeinen vaihe ( final phase) on
FXIII (” tekijän 13″) aiheuttama
palautumattoman hyytymäverkon muodostus.
JOS KOAGULAATIO-puolta ei olisi kontrolloimassa myös sen kehkeytymisen eräässä vaiheessa alkava heräävä vastavaikuttava ANTIKOAGULATIIVINEN puoli ja FIBRINOLYYSI jokainen ihminen koaguloituisi parissa minuutissa
Mutta jos antikoagulaatio ja fibrinolyysi eivät olisi taas puolestaan jarrutettavissa ja hieno. säädettävissä, ihmiskunta menehtyisi verenvuotoon pikaisesti.
Tämän takia koagulaatiopuoli on melko voimakas ja helposti korostuva, jos ihminen joutuu johonkin poikkeustilaan, sillä elinaikaa pidentää globaalisti ottaen hyperkoagulaatiokyky enemmän kuin vuototilan luomiskyky.
KOAGULAATION kaikkein tärkein fysiologinen INHIBIITTORI on nimeltään
ANTITROMBIINI. Se pystyy myös inhiboimaan muita aktivoituneita seriiniproteaaseja kuten TF/VIIa kompleksia, FIXa, FXa, FXIIa.
Toinen ”negatiivisen koagulaatiokontrollin” järjestelmä on
C- ja S-proteiinit, joka voi aktivoitua kun valmistunut trombiini alkaa vaikuttaa trombomoduliiniin (TM) ja niihin struktuurinmuutosta.
Fysiologinen antikoagulaatio järjestelmä käynnistyy asteittain hierarkisessa järjestyksessä trombiinin muodostuttua ja siinä on osatekijänä vaurion lähellä olevan
ehjän endoteelin trombomoduliini TM, johon kiinnittynyt trombiini restrukturoituu ja sitten
aktivoi C-proteiinin S-proteiini kofaktorina ja muodostuu APC,
aktiivi C-proteiini, joka saa hillittyä ja jarrutettua trombiinin kiihtyneen muodostumisen. Se on antikoagulatiivinen.
Tämän APC- funktion kehkeytymsessä voi geneettisillä tekijöillä olla merkitystä, koska koagulaatio-antikoagulaatio- fibrinolyysi- antifibrinolyysi ovat tarkasti ja monivivahteisesti ajoittuvia, säätyviä:
ajoituksen muutoksia voi tapahtua, jos jokin aminohappo on mutatoitunut entsyymiproteiinissa. Koska tekijöitä kuitenkin on tavattoman monta, kompensoitumista tapahtuu ja tulos voi olla “normaali” monista lähtökohdista käsin.
Tiedon lisääntyessä näistä voidaan myös lääkkeelläkin tasapainottaa tilannetta tarvittaessa.
(Huom. lisäykseni: toisaalta kun trombiini vaikuttaa TM:n kanssa
antikoagulatiivisen APC:n se toisaalta taas
saa TAFI:n aktivoitumaan ja TAFIa hillitsee fibrinolyysin voimakkuutta ja tavallaan koettaa pidentään hyytymän olemassaoloa).
- Tapahtumien AJOITUS SEIKKA:
FVL_ mutaatiossa, jota tutkija tarkasti pohti, on kyse aktiivin C-proteiinin (APC)- tekemästä FVa tekijän pilkkomisesta. Tuloksena pitäisi olla inhiboitunut FVa ( FVia).
http://www.scripps.edu/mem/gale/research_files/image009.gif
Pilkkoutuminen tapahtuu normaalisti nopeasti ja täysin: kahteen arginiiniin (506 ja 306). Mutta jos S-proteiinissa on poikkeavuus,
pilkkoutumisen ajoittuminen on hieman poikkeava,
hidastunut. Arg 506 kohta pilkkoutuu nopeasti mutta vain osittain , Arg306 kohta pilkkoutuu täysin, mutta
hitaasti. Siis tämä trombiinin muodostuksen toinen kiihdyttäjä (accelerator) ei jarrutu nopeasti ja täysin, mistä seuraa tiettyä enempää hyperkoagulabiliteettia, "normaalifunktion perässä laahaamista". Siis APC siis kyllä muodostuu, mutta sen teho ei ole paras mahdollinen.
Tekijä mainitsee muistakin hemostaasi tekijöistä seuraavaa.
- ANTITROMBIININ määrä sinänsä ei vaihtele raskauden aikana kuten ei C-proteiinin määrä sinänsä.
- TROMBOSYYTTIEN aktiivisuus nousee raskauden aikana, mutta raskauden viimeisen kolmanneksen aikana niiden lukumäärä laskee lisääntyneen konsumption takia.
- Kudostekijälle (TF) on oma inhibiittorinsa TFPI, jonka määrä nousee raskauden aikana.http://www.med.unc.edu/mackmanlab
- Z-proteaasi-inhiboi tekijää FXa ja proteiini Z lisää reaktiota.
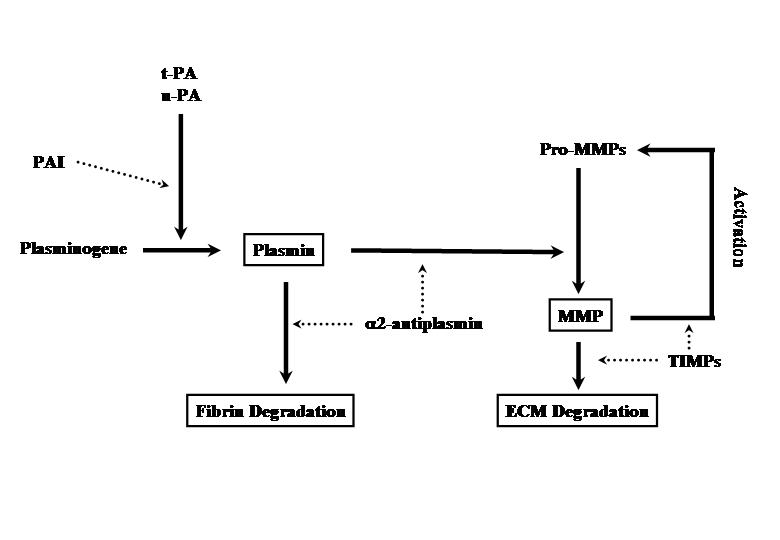
- Entä sitten kun FIBRIINI on jo muodostunut?Fibriini taas aiheuttaa fibrinolyysin aktivoitumisen. Fibriini on proentsyymin ja aktivaattorin asettumisalusta.
- PLASMINOGEENI (plg) on proentsyymi, joka pilkkoutuu vahvaksi PLASMIINI-entsyymiksi(plm).
- t-PA on plasminogeenin aktivaattori, joka e.m. aktivoimisen tekee. Plasminogeeniä sitoutuu fibriiniin ja fibriini itsekin stimuloi tätä pilkkoutumista plasmiiniksi.
- PLASMIINI pilkkoo fibriinistä degradaatiotuotteita (FDP), ja nämä tuotteet sinänsä toimivat luonnollisina antikoagulantteina estäen trombiinivaikutusta ja estäen fibriinin polymeroitumista: ”Natural anticoagulants”.
Mitkä ovat FIBRINOLYYSIN PÄÄESTÄJÄT, inhibiittorit ?
- PAI-1
- alfa2-antiplasmiini
- lipoproteiini (a)
- TAFI = Thrombin Activated Fibrinolysis inhibitor (TAFI :lla ei ole omaa estäjäänä).
http://upload.wikimedia.org/wikipedia/commons/0/0e/Fibrinolysis.png
Kun KOAGULAATIOTEKIJÄ F XIII tekijä on vaikuttanut fibriiniin ja siinä on muodostunut poikkisiltoja, tuloksena syntynyt
palautumaton hyytymä on paljon resistentimpi plasmiinin entsymaattiselle vaikutukselle ja pilkkomisille kuin ennen “ tekijää 13.” ilmenevä fibriinimuoto.
MONIA MUITA TEKIJÖITÄ osallistuu tähän hemohomeostaasiin,
toiset kiihdyttävät
koagulaatiota,
toiset jarruttavat,
toiset kiihdyttävät
antikoagulaatiota
ja toiset jarruttavat,
toiset kiihdyttävät
fibrinolyysiä
ja toiset jarruttavat,
joten vikatoimintojen mahdollisia kohtia on paljon, mutta kukin voi edistää omaa reologista terveyttään ottamalla huomioon terveellisestä elintavasta annetut ohjeet ( liikunta ja ravinto).
Mainittakoon tässä että
plasmiini pystyy aktivoimaan myös matrixmetalloproteaasien (MMPs) (
kudosta sorvaavien) entsyymien joukkoa, joten tulpanjälkeinen kudosten korjaantuminen ja verisuonen avautuminen ennalleen putkeksi on mahdollista tässä yhteistyössä.
Tässä tosin on
kahden alueen sorvausta, riippuen aivan verenkierron tilanteesta-
: verisuonten sisäisestä ( veren fluiditeetistä, rheologisesta funktionaalisuudesta)
tai ulkoisesta tilanteesta ( ECM matrix tapahtumista)
.Verisuonten sisäiseen osallistuu ensisijaisesti plasmiinista itsestään pilkkoutunut
angiostatiini, jota aina muodostuu pikkuisen, mutta tilanteen pahentuessa, sitä generoituu tilanteen mukaisesti- angiogeneesin ja angiostatiinin suhteet rheologiassa pyrkivät tavalla tai toisella olemaan elossapysymistä edistäviä yhteenlaskettuna tuloksena tapahtumista.
u-PA teitse voi plasmiinista käsin aktivoitua MMP-klaskadit ja ne taas vaikuttavat angiostatiinin lisäpilkkoutumista ja ECM tapahtumia. Normaalisti MMP määrät ovat miltei olemattomia kiertävässä veressä, mutta patologisissa tiloissa niitä on kirjavat profiilit taudista riippuen.
Näillä reologisilla ja verisuonia sekä matriksia moduloivilla asioilla on lähinnä merkitystä ulkopuolisen säätelyn tarpeen takia esim
sikiöaikana, reproduktiossa, trauman, raskauden, anestesian ja operaation yhteydessä , mutta aina ne fysiologisissa puitteissa vastaavat kehon jatkuvasta uudistumisesta ja plastisuudesta, normaaliudesta.
http://en.wikipedia.org/wiki/Angiostatin
MMP TIMP kaskadin sijoittuma rheologiseen kaskadiin.
http://www.lbtd.ulg.ac.be/mmp.html
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}