Although
several ADAMs (A disintegrin-like and metalloproteases) have been shown
to contribute to the amyloid precursor protein (APP) metabolism, the
full spectrum of metalloproteases involved in this metabolism remains to
be established. Transcriptomic analyses centred on metalloprotease
genes unraveled a 50% decrease in ADAM30 expression that inversely
correlates with amyloid load in Alzheimer's disease brains. Accordingly,
in vitro down- or up-regulation of ADAM30 expression triggered an
increase/decrease in Aβ peptides levels whereas expression of a
biologically inactive ADAM30 (ADAM30(mut)) did not affect Aβ secretion.
Proteomics/cell-based experiments showed that ADAM30-dependent
regulation of APP metabolism required both cathepsin D (CTSD) activation
and APP sorting to lysosomes. Accordingly, in Alzheimer-like transgenic
mice, neuronal ADAM30 over-expression lowered Aβ42 secretion in neuron
primary cultures, soluble Aβ42 and amyloid plaque load levels in the
brain and concomitantly enhanced CTSD activity and finally rescued long
term potentiation alterations. Our data thus indicate that lowering
ADAM30 expression may favor Aβ production, thereby contributing to
Alzheimer's disease development.
HYAs production has been
observed along the phylogenetic tree, from bacteriophages and other
viruses, pathogenic bacteria, fungi, and invertebrates to vertebrate
animals (26–28).
In vertebrates, different cell types produce these enzymes, and they
are detected in the ECM of diverse organs, including the testis, eyes,
skin, spleen, liver, kidney, and uterus, and in secretions, including
serum, semen and animal venoms (29) (Table 1).
TABLE 1
Table 1 Comparison between HYAL and SVHYA.
HYAs enzymes, also called hyaluronoglucosaminidases, are
members of the class of hydrolases, a subclass of glycosylases (EC
3.2). These enzymes function as glycosidases (EC 3.2.1) due to their
ability to hydrolyze O- and S-glycosyl compounds (30).
HYAs are glycoproteins with a broad range of molecular weights from 7
to 320 kDa. The optimal pH for their action can vary from 3.3 to 7.0 (29).
According to the molecular substrates and products generated by HYAs
enzymatic reactions, these enzymes are classified into three main
subclasses (26–30):
1.
HYAs (EC 3.1.2.35): This subclass includes hyaluronoglucosaminidases
present in semen, serum, tissues, and lysosomes, as well as in
hymenopteran and snake venoms. They possess transglycosidase and
hydrolytic activities. Among the substrates of these enzymes, hyaluronan
is highlighted. In addition, these enzymes act on chondroitin sulfate A
and C and to a lesser extent on dermatan sulfate (chondroitin sulfate
B) and β-heparin. The main product of their catalytic activity is the
tetrasaccharide GlcUA-GlcNAc-GlcUA-GlcNAc.
2.
HYAs (EC 3.1.2.36): This subclass includes hyaluronoglucuronidases that
hydrolyze hyaluronan, resulting in the release of tetra- and
hexasaccharides. These enzymes have been reported in leeches, parasites,
and crustaceans.
3.
HYAs (EC 4.2.2.21): This HYA group is produced by bacterial species and
is characterized as HA lyases. They degrade HA, dermatan sulfate, and
chondroitin sulfate A and C. These enzymes are called
endo-β-N-acetyl-D-hexosaminidases, which act via β elimination since their catalytic activity generates disaccharides.
The
molecular mechanisms of catalysis and substrate specificity are
dictated by the presence of positional and structural catalytic residues
conserved in the species in which HYAs were identified. The amino acid
residues that characterize this enzymatic class are Glu149, which is important for the catalytic mechanism; the Asp147, Tyr220, Trp341 triad, which is responsible for positioning the carbonyl acetamide group for catalysis; and Tyr265, which is responsible for the HYAs specificity for HA. The replacement of the Tyr265 residue for Cys265 switches HYA specificity to chondroitin (2, 29).
2.2 Snake venom hyaluronidases
The initial data
for the SVHYA were obtained during the 1930s. These studies showed that
venoms contained a spreading factor that was able to increase tissue and
blood capillary permeability to Indian ink and to pathogenic bacterial
species. Some authors postulated that this factor would be important for
venom absorption by prey and human victims (35, 36).
In subsequent decades, the presence of spreading factors involved in
efficient toxin delivery was ubiquitously detected in snake venoms.
These factors, which include snake venom metalloproteinases and SVHYA,
are important factors in tissue destruction since their actions are
responsible for ECM breakdown (2, 29).
SVHYA potentiate hemorrhaging, swelling, muscle damage and lethal
effects of purified venom toxins, since its inhibition by monoclonal
antibodies and plant derivative inhibitors substantially decreased the
toxic effects of the venoms (1, 31–34). Thus, based on the available data, SVHYA are considered the main snake venom spreading factors.
Similar
to HYAL, SVHYA are glycoproteins; however, their molecular weight
ranges from 33 to 110 kDa, and they are generally produced as single
chain polypeptides (29). In addition, more than one isoform has been reported in some venoms (1, 31). Harrison and colleagues (2)
scrutinizing cDNA libraries and protein sequences showed that SVHYA
conserve positional and structural catalytic residues that characterize
this enzyme group.
Although hyaluronidases are
ubiquitously expressed in snake venoms, the mechanisms involved in their
effect on HA, which is present in the ECM and bloodstream, and the
inflammatory consequences of these actions are underexplored.
Biochemical studies examining the structure and activity of SVHYA
clustered these enzymes in the EC 3.2.1.35 subclass together with HYAL (2, 33), which were previously shown to trigger inflammatory events (3, 6–10).
Additionally, like HYAL, SVHYA act on HA to generate tetra- and
hexasaccharides, suggesting that they potentially exert
immunopathological effects.
(Sitaatti otettu 8.5. 2023 tähän MMP blogiin, sillä kyynmyrkky on hyvin monen entsyymin seos, jossa vaikutukset eivät ole ainoastaan myrkyn metalloproteinaaseista SVMPs ja sen takia on aiheellista mainita muitakin myrkyllisyyden tekijöitä yhteydessä. Hyaluronaasien osuutena on myrkyn leviämisen edistäminen. Metalloproteinaasit hajoittavat basaalilaminaa. trombiinikaltaiset SVTLEs vaikuttvat veren reologian puolella. Kiniiniä vapauttavat vaikuttavat mikrovaskulaariseen permeabiliteettiin, turvotuksen lisäämiseen , hypovolemiaan. Lektiinit , PLA2 vaikuttavat mastsoluihin ja hepariinin vapautumiseen. Lihastoksinen PLA2 tekee myonekroosia , kehittyy endoteeliperäisiä sytokiineja, trombosytopeniaa havaitaan,, erytrosyyteissä muutoksia, leukosyyttejä verisuoniston ulkopuolelle -rheologisia ilmiöitä on laidasta laitaan- , vuotoa, trombeja, DIC- hypofyysi vaikuttuu, stressihormonit vaikuttuvat, verenmuodostus vaikuttuu. käärmeenpurema on aina otettava vakavasti- joskus tapahtumat ovat hiipiviä ja yllättäen tapahtuu pahenema-
Kuljetus hoitoon ja observaatio ovat tärkeät alkuvaiheet, että lääkärikunta pääsee tapahtumista ajoissa jyvälle.
Kuuma kesä on jälleen alussa ja käärmeet heräävät. .
Käärmeenmyrkyssä voi olla reniinin kaltaisuutta siten että se voi pilkkoa RaAS- järjestelmää spesifisesti siten että vaikutukset järjestelmässä ovat raflaavia; Snake Venom Aspartic proteases SVAPs
Three aspartic proteases (SVAPs) have been isolated from venom of the saw-scaled viper, Echis ocellatus. In confirmation of prior transcriptomic predictions, all three forms match to sequences of either of the two SVAP transcripts (EOC00051 and EOC00123), have a molecular weight of 42 kDa and possess a single N-glycan. The SVAPs act in a renin-like manner,specifically cleaving human and porcine angiotensinogen into angiotensin-1 and possess no general protease activity. Their activity is completely inhibited by the aspartyl protease inhibitorPepstatin A.
1.1 Geographical distribution and dietary acquisition of snakes
Venomous snakes occupy virtually all ecological niches (Vidal et al., 2007). Snakes such as Bitis arietans, Bitis. gabonica, Echis leucogaster, Echis ocellatus, Naja haje, Naja nigricollis, Naja melanoleuca, Dendroaspis jamesoni, Dendroaspis polylepis and Dendroaspis viridis are abundant in tropical and SSA (Tasoulis & Isbister, 2017). As shown in Fig. 1, these snakes occupy different regions of the African continent.
Analysis
of the sequence alignment and the overall three-dimensional structural
properties of Snake venom thrombin-like enzymes (SVTLEs)
Structural comparison among all SVTLEs
Glycosylation and its role in these enzymes
Structure based catalytic mechanisms, processing and inhibition
Structural comparsion between snake venom serine proteinases and SVTLEs
Abstract
Snake venom thrombin-like enzymes
(SVTLEs) constitute the major portion (10–24%) of snake venom and these
are the second most abundant enzymes present in the crude venom. During
envenomation, these enzymes had shown prominently the various
pathological effects, such as disturbance in hemostatic system, fibrinogenolysis, fibrinolysis, platelet aggregation, thrombosis, neurologic disorders, activation of coagulation factors,
coagulant, procoagulant etc. These enzymes also been used as a
therapeutic agent for the treatment of various diseases such as
congestive heart failure, ischemic stroke, thrombotic disorders etc.
Although the crystal structures of five SVTLEs are available in the Protein Data Bank
(PDB), there is no single article present in the literature that has
described all of them. The current work describes the structural
aspects, structure-based mechanism of action, processing and inhibition
of these enzymes. The sequence analysis indicates that these enzymes
show a high sequence identity (57–85%) with each other and low sequence
identity with trypsin (36–43%), human alpha-thrombin (29–36%) and other
snake venom serineproteinases (57–85%). Three-dimensional structural analysis indicates that the loops surrounding the active site are variable both in amino acids composition and length that may convey variable substrate specificity to these enzymes. The surface charge distributions also vary in these enzymes. Docking analysis with suramin
shows that this inhibitor preferably binds to the C-terminal region of
these enzymes and causes the destabilization of their three-dimensional
structure.
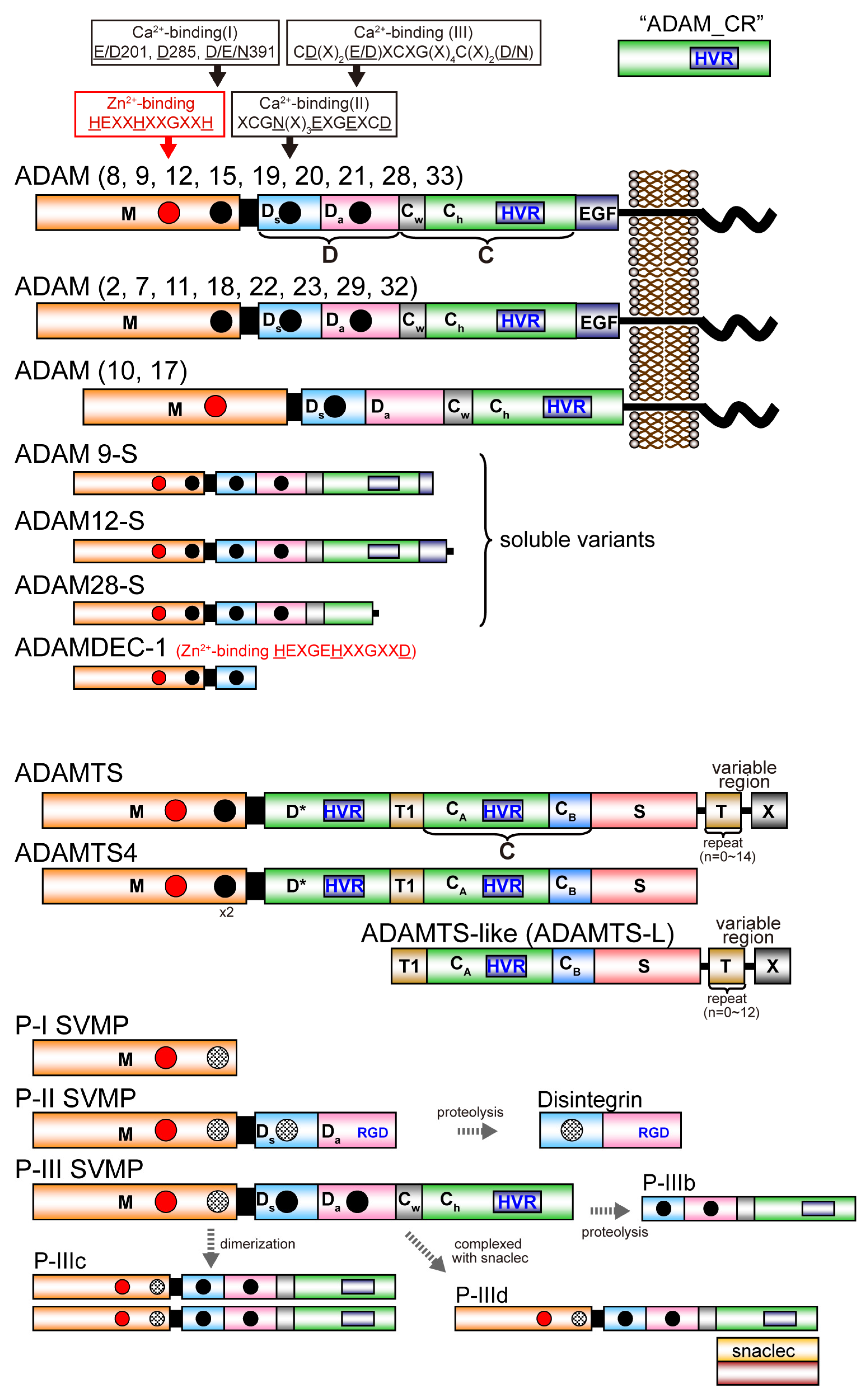
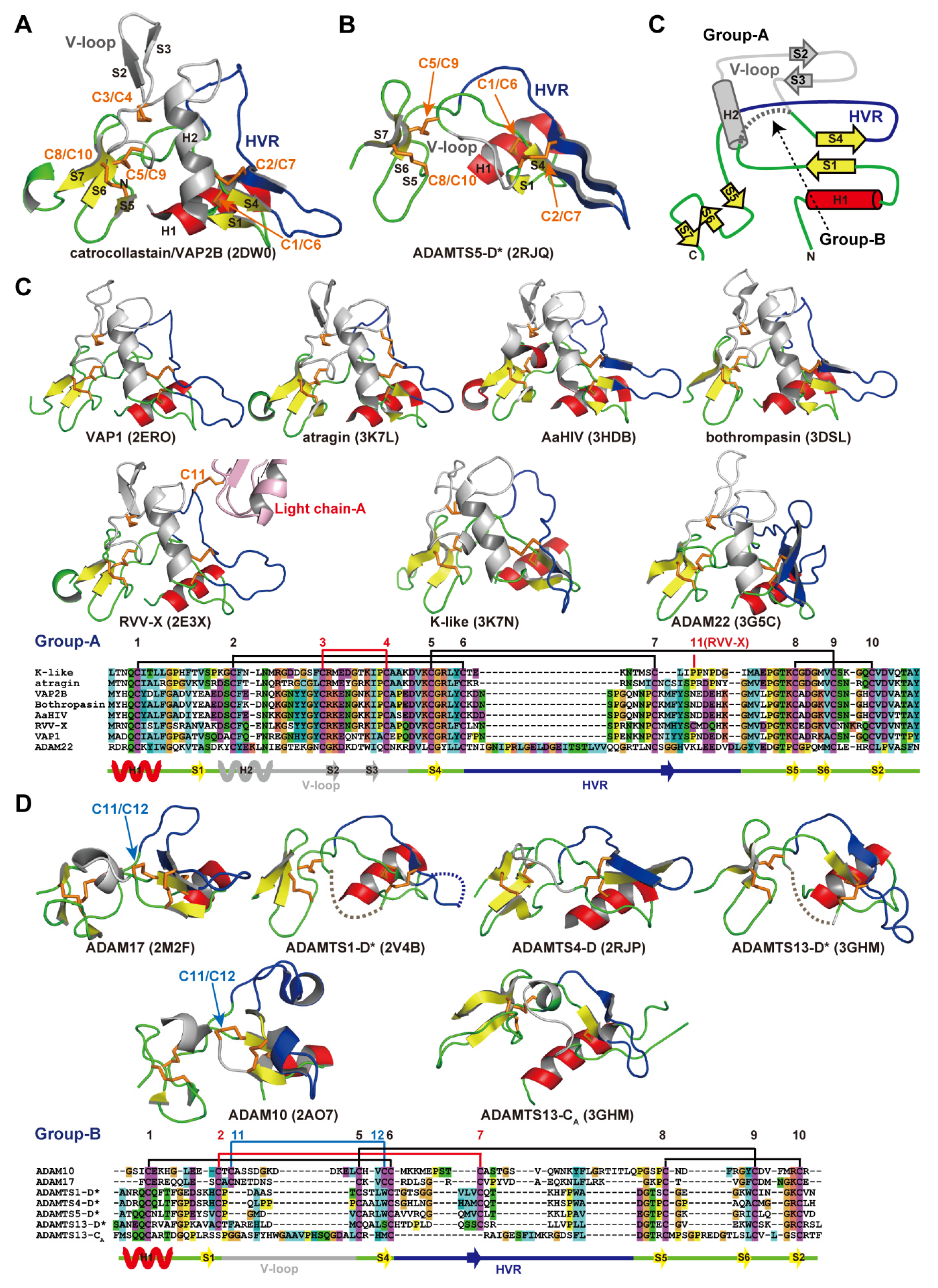
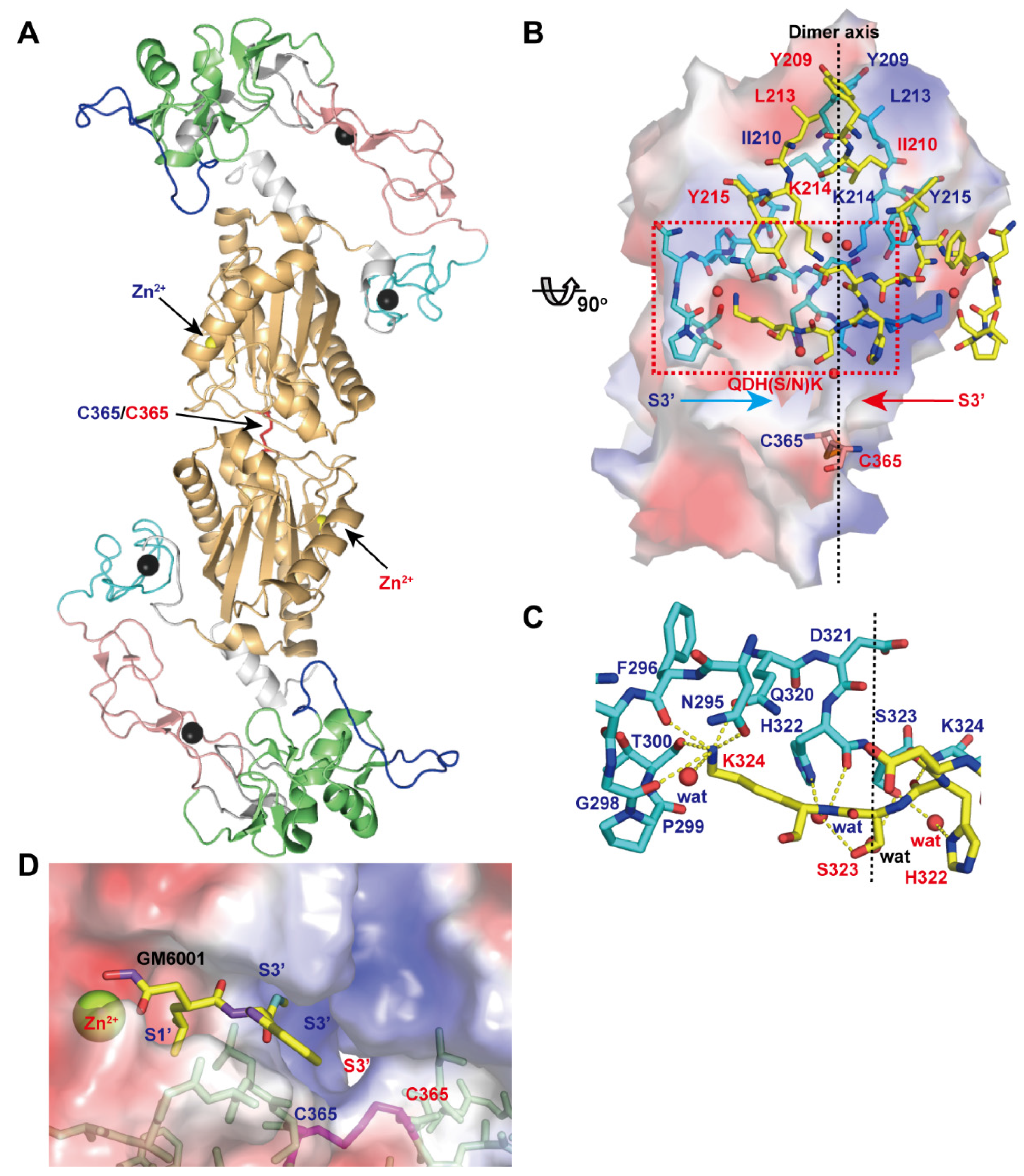
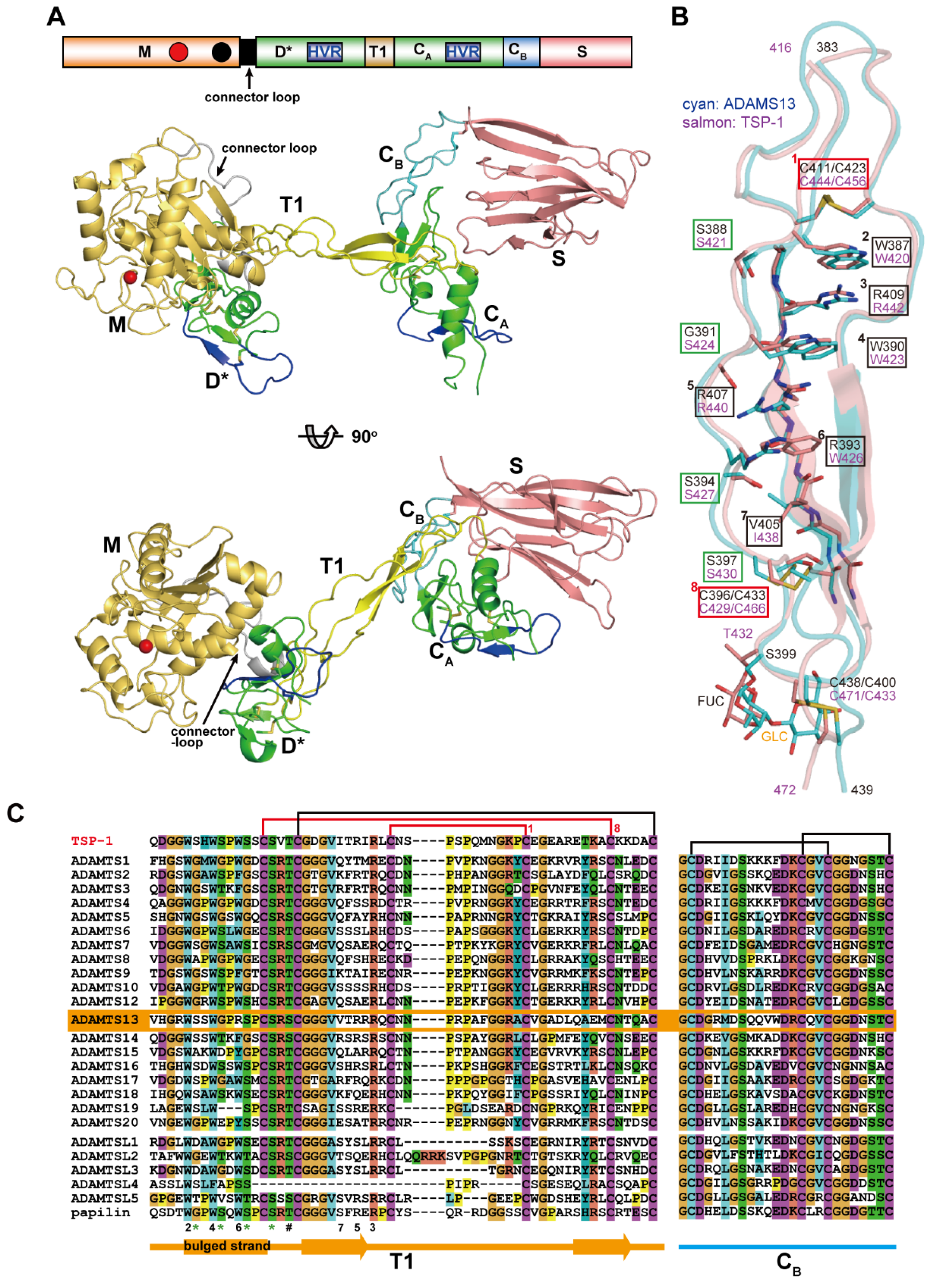
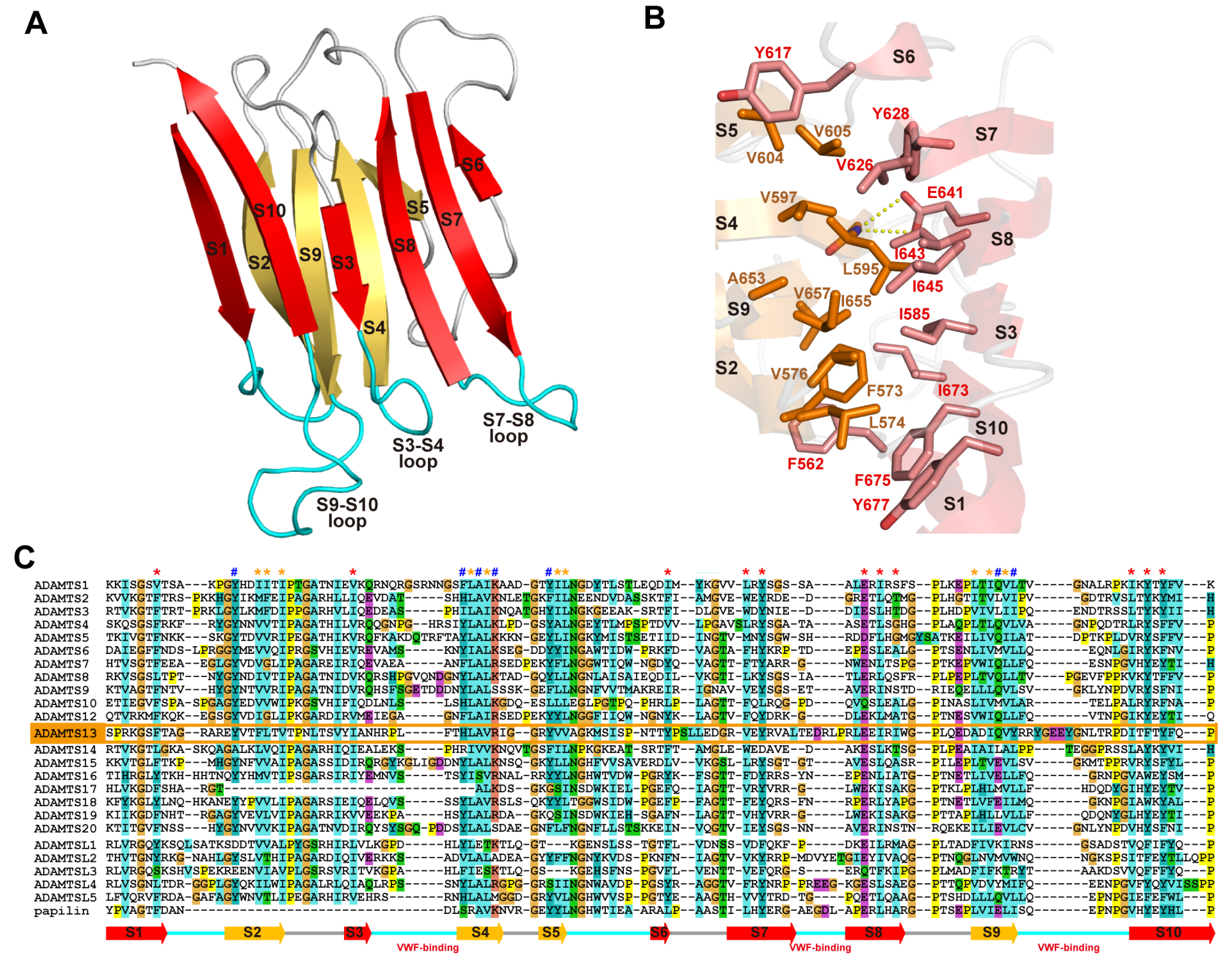
A disintegrin and metalloproteinase (ADAM) family
proteins constitute a major class of membrane-anchored multidomain
proteinases that are responsible for the shedding of cell-surface
protein ectodomains, including the latent forms of growth factors,
cytokines, receptors and other molecules. Snake venom metalloproteinases
(SVMPs) are major components in most viper venoms. SVMPs are primarily
responsible for hemorrhagic activity and may also interfere with the
hemostatic system in envenomed animals.
SVMPs are phylogenetically most
closely related to ADAMs and,
together with ADAMs and related ADAM with
thrombospondin motifs (ADAMTS) family proteinases,
constitute
adamalysins/reprolysins or the M12B clan (MEROPS database) of
metalloproteinases.
Although the catalytic domain structure is
topologically similar to that of other metalloproteinases such as matrix
metalloproteinases, the M12B proteinases have a modular structure with
multiple non-catalytic ancillary domains that are not found in other
proteinases. Notably, crystallographic studies revealed that, in
addition to the conserved metalloproteinase domain, M12B members share a
hallmark cysteine-rich domain designated as the “ADAM_CR” domain.
Despite their name, ADAMTSs lack disintegrin-like structures and instead
comprise two ADAM_CR domains.
This review highlights the current state
of our knowledge on the three-dimensional structures of M12B
proteinases, focusing on their unique domains that may collaboratively
participate in directing these proteinases to specific substrates.
Snake venom is a rich source of peptides and proteins with a wide range
of actions. Many of the venom components are currently being tested for
their usefulness in the treatment of many diseases ranging from
neurological and cardiovascular to cancer. It is also important to
constantly search for new proteins and peptides with properties not yet
described.
The venom of Vipera berus berus has hemolytic, proteolytic and cytotoxic properties, but its exact
composition and the factors responsible for these properties are not
known. Therefore, an attempt was made to identify proteins and peptides
derived from this species venom by using high resolution two-dimensional
electrophoresis and MALDI ToF/ToF mass spectrometry.
A total of 11
protein classes have been identified mainly proteases but also l-amino acid oxidases, C-type lectin like proteins, cysteine-rich venom proteins and phospholipases A2
and 4 peptides of molecular weight less than 1500 Da.
Most of the
identified proteins are responsible for the highly hemotoxic properties
of the venom. Presence of venom phospholipases A2 and l-amino
acid oxidases cause moderate neuro-, myo- and cytotoxicity. All
successfully identified peptides belong to the bradykinin-potentiating
peptides family.
The mass spectrometry data are available via
ProteomeXchange with identifier PXD004958.
1, Introduction
Venom
is a complex mixture of various chemicals that are used to kill or
immobilize the victim and eventually help digestion. These substances
affect nervous, muscular and cardiovascular systems. Most of the toxic
substances, as much as 95%, contained in the venom of snakes are
polypeptides: enzymes and non-enzymatic proteins. Depending on the
genus, snakes produce venom of different composition and mechanisms of
action, but within the family it has similar composition [1].
Vipera berus berus
or common European adder is found in Europe and Asia in the areas of
wetlands, peat bogs and forests, where they can find sunny slopes and
glades. Depending on the area in which an individual resides, coloration
varies from gray, blue-gray, brown, green-brown, red-brown to black. On
the back, a distinctive dark zigzag is present, and on its head a dark
stain in the shape of the letter H, V or X. The head is clearly
separated from the trunk, triangular, flattened, and covered with tiny
plates [2].
Venom
of the common European adder is a yellow liquid consisting of
approximately 25 proteins and peptides with enzymatic activity. Total
composition of it is not fully known. Venom ingredients immobilize the
victim and initialize digestion of the tissue near the site of the bite.
The venom has hemolytic, proteolytic and cytotoxic properties. It
consists of: protease, phospholipase, hyaluronidase, metalloproteinases,
phosphodiesterases and l-amino acid oxidase. The presence of these families of compounds cause edema, disruption of homeostasis and hypovolemia [3,4].
Only a few of venom components are described in the V. berus berus
species. Presence of the most of the venom components is inferred from
the properties of the venom itself. Currently venom of many snakes is
intensively studied because of the huge variety of proteins that occur
there. Knowledge of the venom proteome and biological properties of the
individual components may constitute a valuable source of new drugs.
Collected information might also help in new drug design for use in the
treatment of cardiovascular diseases, nervous system disorders, or
cancer [1].
The aim of the study was to determine the composition of venom protein and peptide produced by adult V. berus berus and it is the first such a full proteomic description for this species.
2. Results
2.1. Proteome
The combined venom from adult Vipera berus berus
individuals (male and female) was separated by two-dimensional
electrophoresis in two pH ranges, 3–10 and 5–8. From the obtained
polyacrylamide gels all visible spots were cut out, and then subjected
to tryptic digestion procedure. All samples were analyzed by mass
spectrometry MALDI ToF/ToF. Polyacrylamide gels show that the most
proteins of this venom are concentrated in the pH 5–8, and only a few,
having a molecular weight below 20 kDa, fall outside the above range of
pH (Figure 1 and Figure 2).
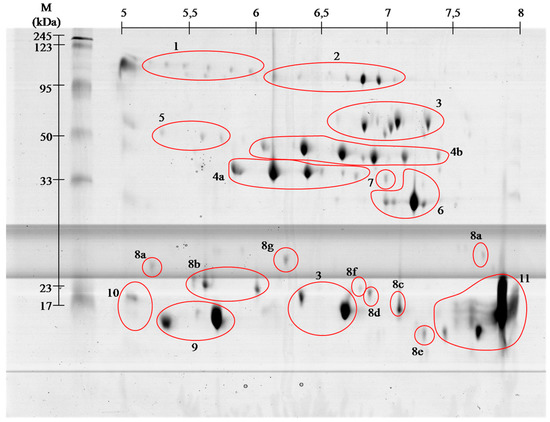
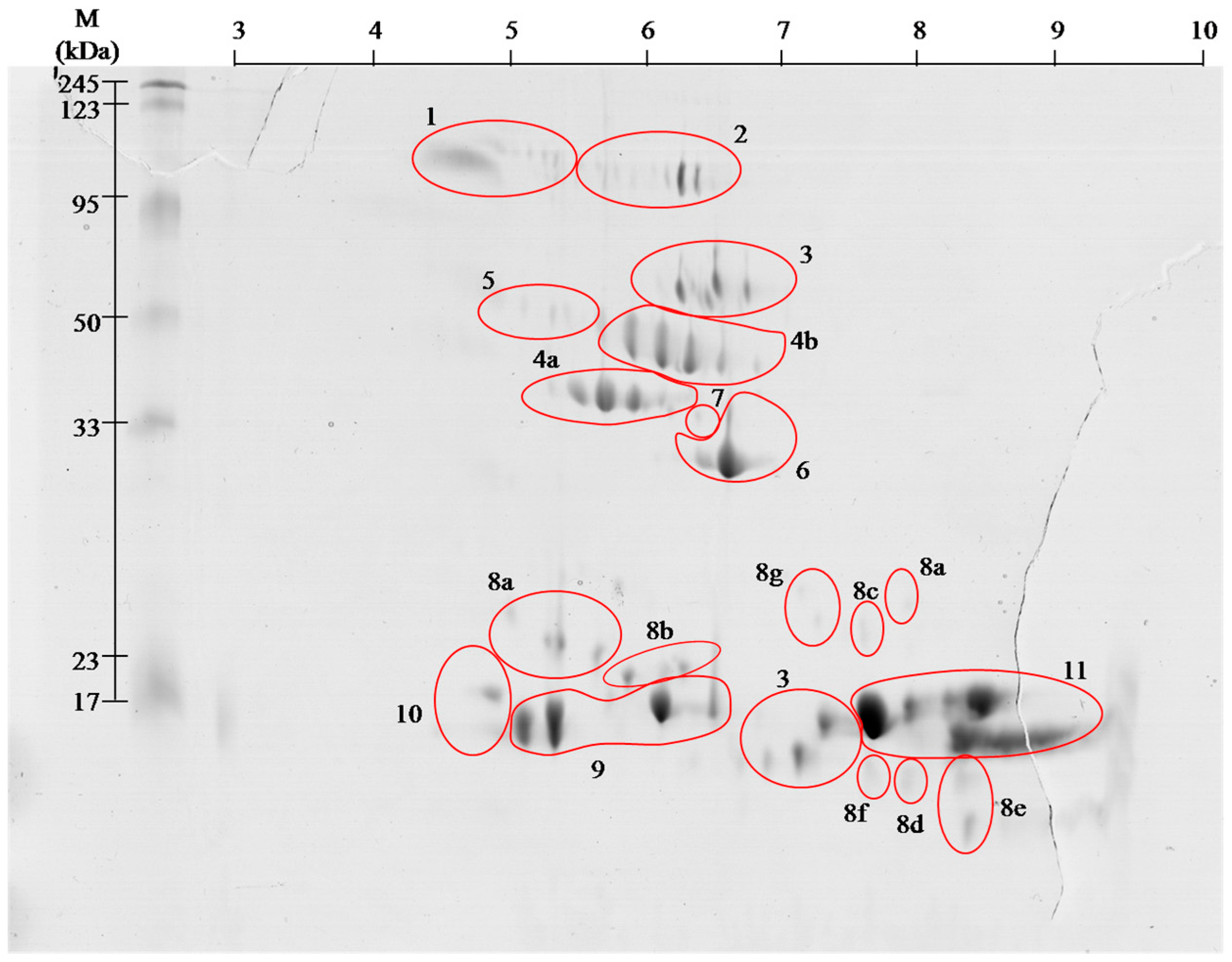
Figure 1.
Representative 2-D protein map in 3–10 pH range, obtained from V. berus berus venom with identified protein groups shown: 1, Angiotensin-like peptide; 2, Metalloproteinase H3; 3, l-amino acid oxidase; 4, Serine proteases: (a) VLSp and (b) nikobin; 5, Beta-fibrogenase brevinase; 6, Cysteine rich venom protein; 7, Snake venom metalloproteinasesSV-s ; 8, Snaclec: (a) rhinocetin, (b) snaclec 14, (c) snaclec B6, (d) echicetin, (e) snaclec 1, (f) rhodocetin/A13, and (g) jerdonibitin; 9, Acidic phospholipases; 10, Basic phospholipases; and 11,
Neutral phospholipase. The proteins were separated by
isoelectrofocusing at pH range 3–10, then distributed on polyacrylamide
gels by SDS-PAGE and stained with colloidal Coomassie Brilliant Blue
G-250. Molecular weight (MW) and pH 3–10 scale are shown.
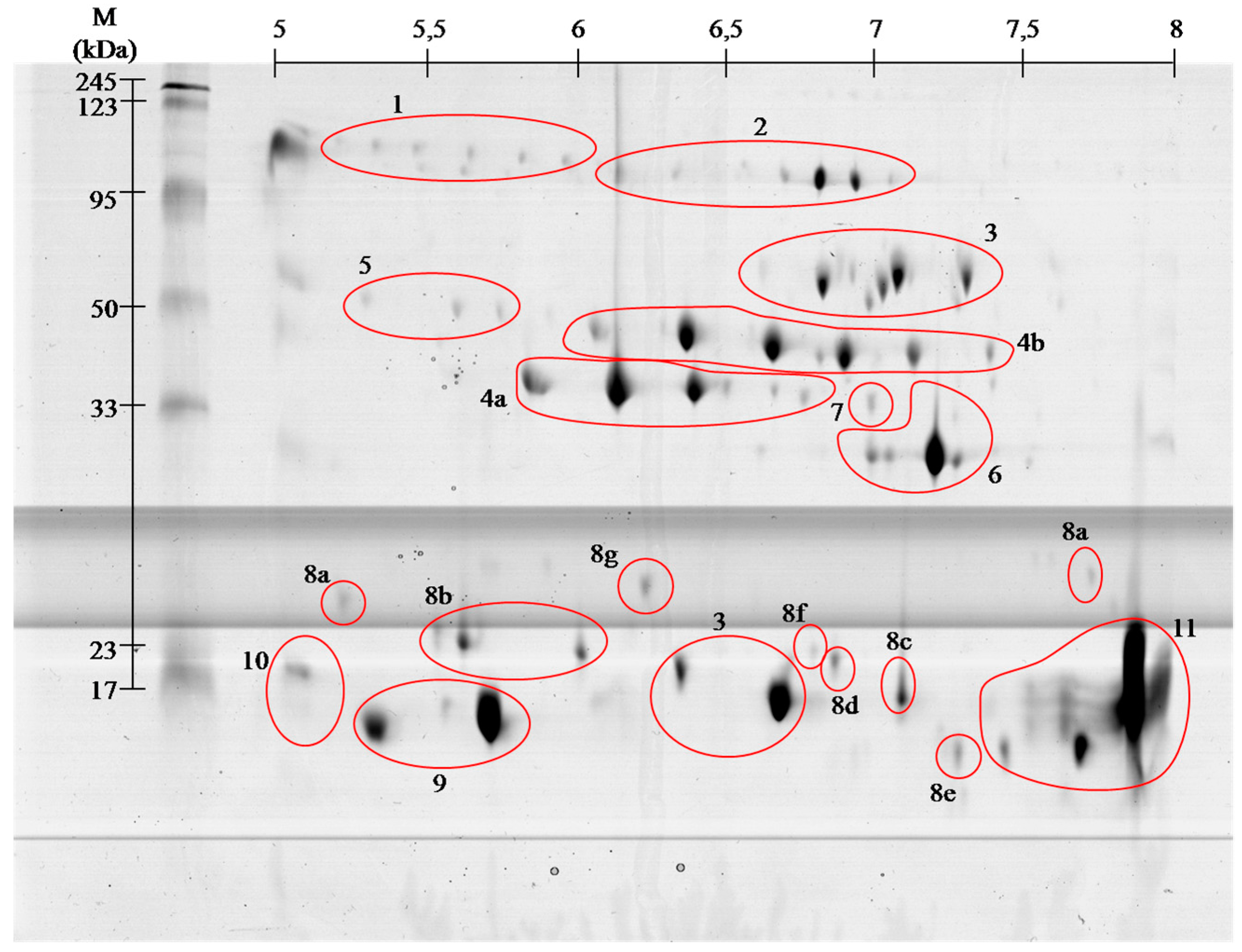
Figure 2.Representative 2-D protein map in 5–8 pH range, obtained from V. berus berus venom with identified protein groups shown: 1, Angiotensin-like peptide; 2, Metalloproteinase H3; 3, l-amino acid oxidase; 4, Serine proteases: (a) VLSp and (b) nikobin; 5, Beta-fibrogenase brevinase; 6, Cysteine rich venom protein; 7, Snake venom metalloproteinases; 8, Snaclec: (a) rhinocetin, (b) snaclec 14, (c) snaclec B6, (d) echicetin, (e) snaclec 1, (f) rhodocetin/A13, and (g) jerdonibitin; 9, Acidic phospholipases; 10, Basic phospholipases; and 11,
Neutral phospholipase. The proteins were separated by
isoelectrofocusing at pH range 3–10, then distributed on polyacrylamide
gels by SDS-PAGE and stained with colloidal Coomassie Brilliant Blue
G-250. Molecular weight (MW) and pH 3–10 scale are shown.
On the basis of performed identification, proteins
have been grouped according to their class. Proteins were grouped by
combining the results of both pH ranges of 3–10 (Figure 1), and 5–8 (Figure 2)
separation. The numbers on gels correspond to the different classes of
proteins. On the gels with broader range of pH, proteins having an
isoelectric point above pH 8 can be seen, whereas no proteins were
observed in pH below 4.
Complete list of identified proteins is summarized in Table 1. Representative MS and MS/MS spectra for all identified proteins and peptides have been included as Supplementary Materials.
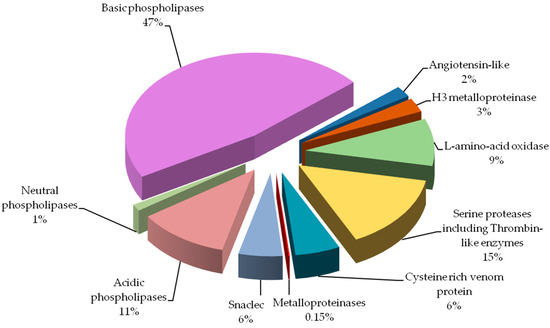
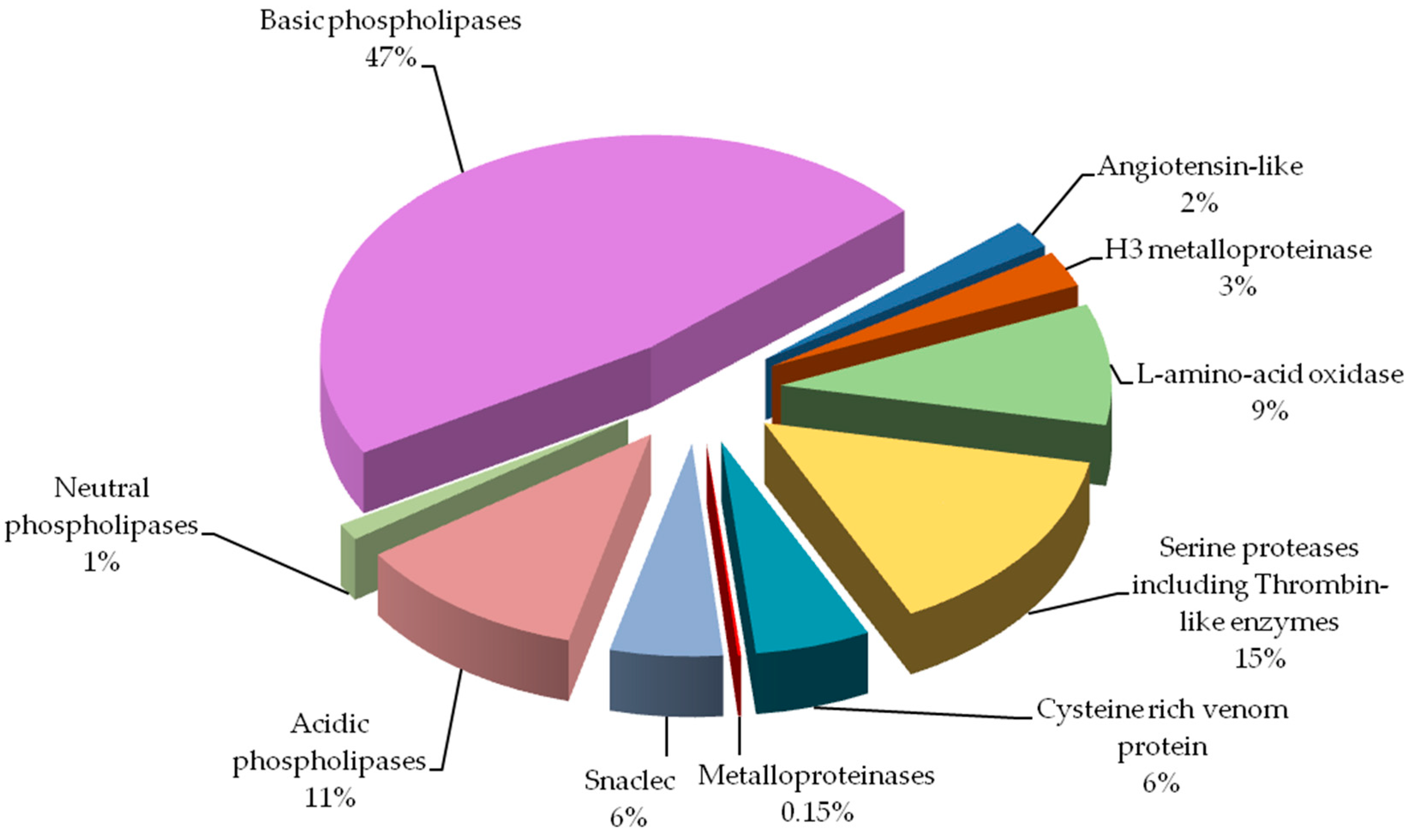
Percentage of protein groups in Vipera berus berus venom is presented in Figure 3.
By far the largest share of the analyzed venom are phospholipases
(almost 60%). Other groups containing a significant amount of protein
are: serine proteases and l-amino-acid oxidase. Angiotensin-like potential protein and metalloproteinases have been detected in the lowest amounts.
Table 1.
Composition of V. berus berus venom proteins.
Snake venom is a rich source of peptides and
proteins with a wide range of actions. Many of the venom components are
currently being tested for their usefulness in the treatment of many
diseases ranging from neurological and cardiovascular to cancer. It is
also important to constantly search for new proteins and peptides with
properties not yet described. The venom of Vipera berus berus
has hemolytic, proteolytic and cytotoxic properties, but its exact
composition and the factors responsible for these properties are not
known. Therefore, an attempt was made to identify proteins and peptides
derived from this species venom by using high resolution two-dimensional
electrophoresis and MALDI ToF/ToF mass spectrometry. A total of 11
protein classes have been identified mainly proteases but also l-amino acid oxidases, C-type lectin like proteins, cysteine-rich venom proteins and phospholipases A2
and 4 peptides of molecular weight less than 1500 Da. Most of the
identified proteins are responsible for the highly hemotoxic properties
of the venom. Presence of venom phospholipases A2 and l-amino
acid oxidases cause moderate neuro-, myo- and cytotoxicity. All
successfully identified peptides belong to the bradykinin-potentiating
peptides family. The mass spectrometry data are available via
ProteomeXchange with identifier PXD004958.
Venom
is a complex mixture of various chemicals that are used to kill or
immobilize the victim and eventually help digestion. These substances
affect nervous, muscular and cardiovascular systems. Most of the toxic
substances, as much as 95%, contained in the venom of snakes are
polypeptides: enzymes and non-enzymatic proteins. Depending on the
genus, snakes produce venom of different composition and mechanisms of
action, but within the family it has similar composition [1].
Vipera berus berus
or common European adder is found in Europe and Asia in the areas of
wetlands, peat bogs and forests, where they can find sunny slopes and
glades. Depending on the area in which an individual resides, coloration
varies from gray, blue-gray, brown, green-brown, red-brown to black. On
the back, a distinctive dark zigzag is present, and on its head a dark
stain in the shape of the letter H, V or X. The head is clearly
separated from the trunk, triangular, flattened, and covered with tiny
plates [2].
Venom
of the common European adder is a yellow liquid consisting of
approximately 25 proteins and peptides with enzymatic activity. Total
composition of it is not fully known. Venom ingredients immobilize the
victim and initialize digestion of the tissue near the site of the bite.
The venom has hemolytic, proteolytic and cytotoxic properties. It
consists of: protease, phospholipase, hyaluronidase, metalloproteinases,
phosphodiesterases and l-amino acid oxidase. The presence of these families of compounds cause edema, disruption of homeostasis and hypovolemia [3,4].
Only a few of venom components are described in the V. berus berus
species. Presence of the most of the venom components is inferred from
the properties of the venom itself. Currently venom of many snakes is
intensively studied because of the huge variety of proteins that occur
there. Knowledge of the venom proteome and biological properties of the
individual components may constitute a valuable source of new drugs.
Collected information might also help in new drug design for use in the
treatment of cardiovascular diseases, nervous system disorders, or
cancer [1].
The aim of the study was to determine the composition of venom protein and peptide produced by adult V. berus berus and it is the first such a full proteomic description for this species.
2. Results
2.1. Proteome
The combined venom from adult Vipera berus berus
individuals (male and female) was separated by two-dimensional
electrophoresis in two pH ranges, 3–10 and 5–8. From the obtained
polyacrylamide gels all visible spots were cut out, and then subjected
to tryptic digestion procedure. All samples were analyzed by mass
spectrometry MALDI ToF/ToF. Polyacrylamide gels show that the most
proteins of this venom are concentrated in the pH 5–8, and only a few,
having a molecular weight below 20 kDa, fall outside the above range of
pH (Figure 1 and Figure 2).
Figure 1.
Representative 2-D protein map in 3–10 pH range, obtained from V. berus berus venom with identified protein groups shown: 1, Angiotensin-like peptide; 2, Metalloproteinase H3; 3, l-amino acid oxidase; 4, Serine proteases: (a) VLSp and (b) nikobin; 5, Beta-fibrogenase brevinase; 6, Cysteine rich venom protein; 7, Snake venom metalloproteinases; 8, Snaclec: (a) rhinocetin, (b) snaclec 14, (c) snaclec B6, (d) echicetin, (e) snaclec 1, (f) rhodocetin/A13, and (g) jerdonibitin; 9, Acidic phospholipases; 10, Basic phospholipases; and 11,
Neutral phospholipase. The proteins were separated by
isoelectrofocusing at pH range 3–10, then distributed on polyacrylamide
gels by SDS-PAGE and stained with colloidal Coomassie Brilliant Blue
G-250. Molecular weight (MW) and pH 3–10 scale are shown.
Figure 2.
Representative 2-D protein map in 5–8 pH range, obtained from V. berus berus venom with identified protein groups shown: 1, Angiotensin-like peptide; 2, Metalloproteinase H3; 3, l-amino acid oxidase; 4, Serine proteases: (a) VLSp and (b) nikobin; 5, Beta-fibrogenase brevinase; 6, Cysteine rich venom protein; 7, Snake venom metalloproteinases; 8, Snaclec: (a) rhinocetin, (b) snaclec 14, (c) snaclec B6, (d) echicetin, (e) snaclec 1, (f) rhodocetin/A13, and (g) jerdonibitin; 9, Acidic phospholipases; 10, Basic phospholipases; and 11,
Neutral phospholipase. The proteins were separated by
isoelectrofocusing at pH range 3–10, then distributed on polyacrylamide
gels by SDS-PAGE and stained with colloidal Coomassie Brilliant Blue
G-250. Molecular weight (MW) and pH 3–10 scale are shown.
On the basis of performed identification,
proteins have been grouped according to their class. Proteins were
grouped by combining the results of both pH ranges of 3–10 (Figure 1), and 5–8 (Figure 2)
separation. The numbers on gels correspond to the different classes of
proteins. On the gels with broader range of pH, proteins having an
isoelectric point above pH 8 can be seen, whereas no proteins were
observed in pH below 4.
Complete list of identified proteins is summarized in Table 1. Representative MS and MS/MS spectra for all identified proteins and peptides have been included as Supplementary Materials.
Table 1.
Composition of V. berus berus venom proteins.
Percentage of protein groups in Vipera berus berus venom is presented in Figure 3.
By far the largest share of the analyzed venom are phospholipases
(almost 60%). Other groups containing a significant amount of protein
are: serine proteases and l-amino-acid oxidase. Angiotensin-like potential protein and metalloproteinases have been detected in the lowest amounts.
Figure 3.
Protein groups of Vipera berus berus venom. Each group is represented as a percent fraction of the particular protein spots present on the gels.
2.2. Peptidome
Peptides
of less than 3 kDa were obtained by filtration and analyzed directly by
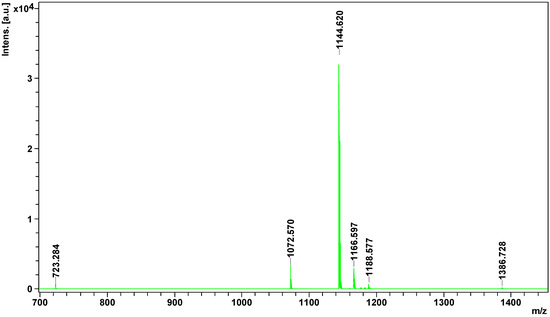
the MALDI ToF/ToF mass spectrometry. In the obtained spectrum eight
signals from the candidate peptides were found, all with apparent mass
less than 1500 Da (Figure 4).
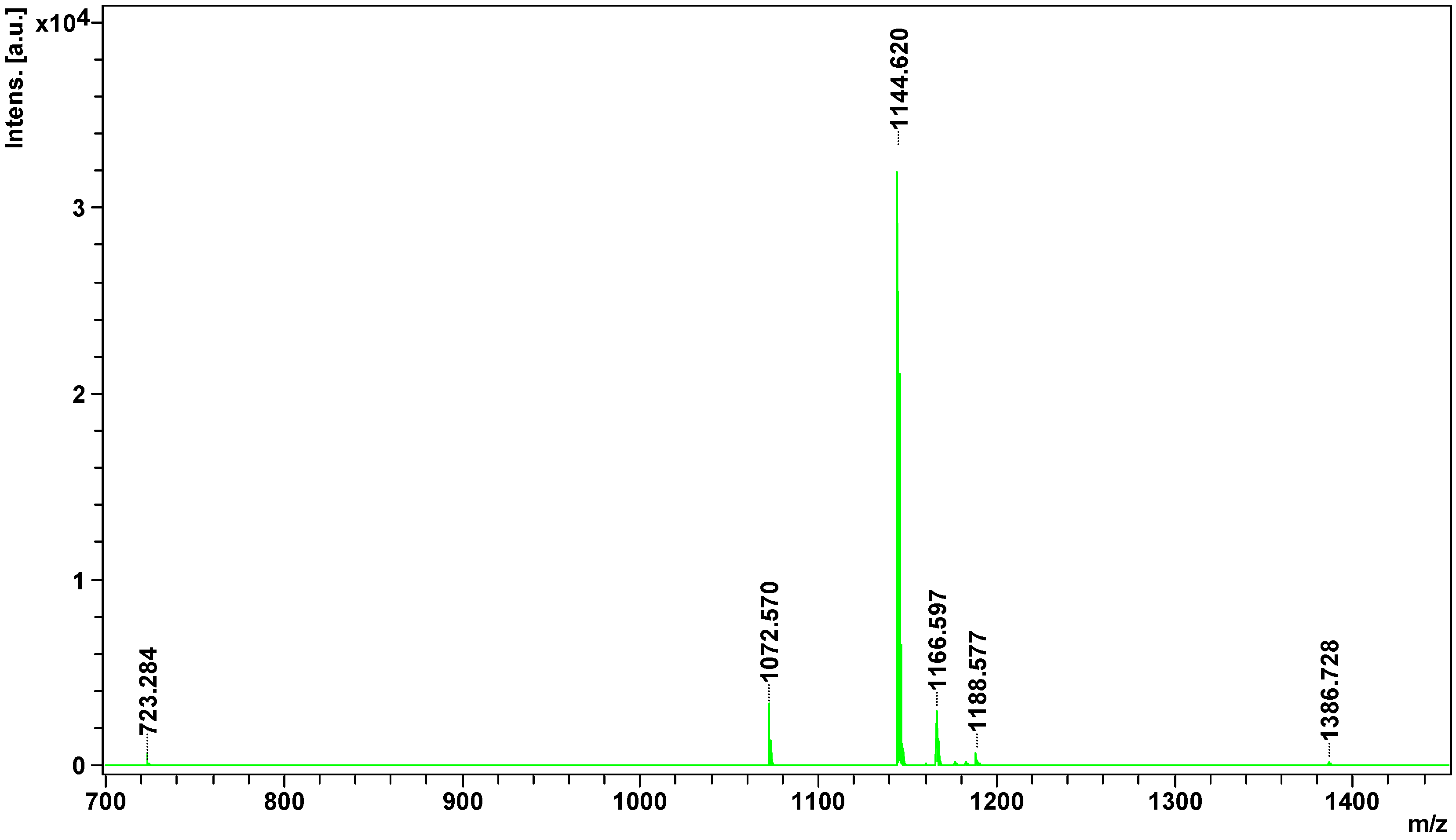
Figure 4.
Mass spectrum of peptidome fraction of Vipera berus berus venom obtained on MALDI ToF/ToF mass spectrometer.
All potential peptides were sequenced in LIFT mode. For parent ion 1386.728 m/z, 178 signals were obtained in the fragmentation spectrum; for 1188.5767 m/z, 140 signals; 1182.557 m/z, 114 signals; for 1176.600 m/z, 109 signals; 1166.597 m/z, 141 signals; 1144.620 m/z, 80 signals; and for 1072.570 m/z, 72 signals. Sequencing of parent ion 723.284 m/z failed. Sequences of four peptides obtained from SwissProt and NCBInr data bases are summarized in Table 2.
Table 2.
Composition of the peptidome of V. berus berus venom.
3. Discussion
Venoms
produced by snakes consist of many components, of which proteins and
peptides are the largest group. Many of these components have a
synergistic effect, which ensures the quick effect of venom on prey.
Victims hunted by a given snake species often belong to different
taxonomic groups, and have developed a variety of safeguards against
bites and its consequences. Therefore, the venom has agents acting
“universally” on a wide range of organisms, as well as those whose
activity is directed against a specific prey molecular targets [5].
For each agent in the human body associated with hemostasis,we may find a homolog, activator or an inhibitor in the venom [6] operating on the principle of protein-protein interactions or enzymatic proteolysis [7].
Hemotoxic venoms, like the one produced by a common European adder,
affect blood vessel walls, platelets, coagulation, anticoagulation and
fibrinolysis. It often happens that, in the venom of a single species we
find components that are antagonists to hemostasis and even to
individual factors associated with it [7].
Venom of V. berus berus consists of approximately 25 proteins and peptides with enzymatic activity [3,4], and the total venom composition only of some Russian specimens has been described so far [8]. Obtained gels of V. berus berus
venom proteins contain even greater number of spots, but the
identification using MALDI ToF/ToF showed that they belong only to 11
families. With high probability it can be assumed that the proteins in
this venom are highly post-translationally modified, as it is shown
clearly by visible spots trains in gels (Figure 1 and Figure 2). This phenomenon is characteristic for Viperidae family and was described already several times [9,10].
Our study indicates that the composition of the analyzed venom differs from that recently described in Latinović et al. [8].
However, direct comparison of obtained results from those two studies
is not possible. Latinović et al. used normalized volumes of
corresponding 1-D gel protein bands and areas of elution peaks from
RP-HPLC for protein abundance estimation. On the other hand our study
employs protein quantity estimation method based on spots volume from
obtained 2-D gels after sample separation. Furthermore, it is not
possible to incorporate our peptidome results in to the protein chart
because of the method we used, as we have identified peptidome directly,
without prior separation. In this case, MALDI ToF/ToF technique does
not provide quantitative data, so we cannot determine the content of
individual peptides in the venom. Keeping that in mind, direct
comparison of the percent shares of major protein groups would indicate
significantly larger amount of phospholipases A2(59% vs.
10%), and much lower amount of serine proteinases (15% vs. 31%) in our
results. The most prominent difference observed would be
metalloproteinases share: 0.15% vs. 19%. Biological explanation for
observed discrepancies would be snake gender, Latinović et al. does not
declare it, and the snake habitat, in our case venom was obtained from
the snakes captured in natural environment in Slovak Republic, or age of
snakes and type of food. Influence of these factors was described
before and could attribute to the observed differences [11].
Vipera berus berus venom has mainly hemotoxic activity and identified proteins clearly meet the criteria for a wide range of hemotoxins [3,4].
Hemotoxins can be classified based on their effects on the following groups [6,7]:
(i) activating blood coagulation factors;
(ii) anticoagulant agents;
(iii) inhibitors and activators of platelets;
(iv) agents affecting
fibrinolysis; and
(v) hemorrhagins.
Proteins of the first group affect
clotting factors or directly coagulate the fibrinogen (thrombin-like
enzymes) (spots # 4 and 5). Most of them, however, cause the formation
of fibrinopeptide A or B or, rarely both as it is in nature. Therefore,
created clots are unstable and prone to endogenous or venom-induced
fibrinolysis, which in turn leads to fibrinolysis syndrome
In the
anti-coagulant agents group, we include those components of the venom,
which inhibit tenas. There are mostly serine proteases (# 4 and 5),
protein C activators and phospholipases A2 (# 9–11).
Platelet
activating proteins cause thrombocytopenia and are predominantly C-type
lectin like proteins (# 8), and thrombin-like enzymes (# 4 and 5). In
turn, deactivation of platelets and following hemorrhage is caused by
disintegrins and snake venom metalloproteases (SVMPs) (# 2 and 7).
The
group of proteins responsible for fibrinolysis includes protein directly
capable of disrupting the fibrin (# 4 and 5) or plasmin activators.
The
last group of proteins is the hemorrhagins–cytolysins damaging blood
vessels and causing hemorrhages. They mostly include metalloproteases (#
2 and 7) [4,5].
We found all the above-described groups of proteins in the venom of Vipera berus berus. The specificity of these proteins clearly explains hemotoxic properties of this venom.
Most
diverse group of proteins in European adder venom is the snaclec
proteins belonging to C-type lectin like proteins. Most snaclec type
proteins are non-enzymatic homodimers of a weight 26 and 28 kDa composed
of subunits with weight about 13 and 18 kDa, and are responsible for
the erythrocytes agglutination. They may also take the form of
heterodimers or oligomers, and contribute to the activation or
inhibition of human platelets [12]. Performed separation under denaturing conditions confirms their monomeric weight in the range of 15 to 25 kDa (Figure 1 and Figure 2). In the V. berus berus venom we identified eight homologues of these proteins from different species of Viperidae (# 8a–8g), constituting 5.5% of venom proteins and this is the first finding of these proteins in this species.
The largest group of proteins identified in the adder venom is a family of phospholipases A2(PLA2) (60%). They are small enzymes with a mass of approximately 14 kDa corresponding to about 115–133 amino acid residues [13].
Depending on the amino acid composition they are divided into acidic,
basic and neutral–all three groups we have identified in European adder
(# 9–11). The snake venom’s phospholipases of group I and II are widely
distributed in many snake species and are important neuro- and miotoxic
agents, causing the immobilization of the prey. Often in the venom of
snakes different types of phospholipases are present, which cause
different pharmacological effects starting with blood coagulation
disorders, through the inhibition of platelet aggregation, to blocking
of neuromuscular signaling and skeletal muscle paralysis [14]. On gels (Figure 1 and Figure 2)
the areas with these proteins appear in the 15 kDa region throughout
the full used pH range, wherein the acidic, basic and neutral
phospholipase were identified, confirming the literature data [13,14].
Serine
proteases, also known as thrombin-like enzymes, are another big
identified group (15%, # 4 and 5). They constitute a collection of
enzymes that catalyze reactions involving a wide range of the blood
coagulation cascade, fibrinolysis and platelet aggregation. Specific
serine proteases catalyze usually only one or a few of the many
reactions involved in blood coagulation. They have the ability to cut
fibrinogen in the way similar to thrombin. This results in clot
formation, acting not only through participation in the coagulation
pathway, but also by direct platelet aggregation [15].
The molecular weight of these enzymes ranges from about 30 to 60 kDa.
On the obtained polyacrylamide gels serine proteases are located in the
area of pH 5–8 and the weight range of 35–50 kDa (Figure 1 and Figure 2).
From the pharmacological point of view this group of proteins is very
interesting and promising since they could be used in
hyperfibrinogenemia treatment, an important risk factor for ischemic
stroke and peripheral artery diseases [16].
In the venom of the Viperidae
family all classes of SVMPs (snake venom metalloproteinases) are
present, playing an important role in immobilizing prey by blocking the
transmission of nerve signals, and tissue proteolysis in the initial
digestion. They play an important role in the impairment of blood
clotting causing immediate local bleeding and delayed internal bleeding [17,18,19].
As it is apparent from this research (# 2 and # 7) SVMPs type III
containing metalloproteinase, disintegrin-like and a cysteine-rich
domains are the largest class of metalloproteinases in V. berus berus
venom. However, this class has a very small share in the venom
proteome, less than 0.5%. Interestingly, earlier studies indicate a much
larger share of this group of proteins in the venom of V. berus berus [8].
In
the upper part of the gel a group of proteins identified as
Angiotensin-like peptide 2 (# 1) was found. The molecular weight of this
peptide is about 1 kDa, and the spot which contain proteins with this
short sequence on the basis of which the identification was made, have
weight almost one hundred times greater. This probably means that in the
venom of European adder there is so far undescribed protein that is
vasoactive, i.e., has a constricting or dilating effect on the caliber
of blood vessels [20].
A second possibility is that the series of spots visible on gels (# 1)
contains the precursors of bioactive peptides. Due to the fact that not
all peptides were identified, there is a chance that considered venom
includes peptides having angiotensin-like properties. However, this
result requires more research as only one short peptide belonging to
this protein was identified.
Besides basic
phospholipase only two other proteins were assied to the database
entries as coming from the examined species. These include l-amino acid oxidases (# 3) and venom cysteine-rich proteins (# 6). l-amino
acid oxidases are present in venoms of many snakes in large quantities
and their toxicity is primarily due to oxidative stress induced by H2O2, which is produced in enzymatic reaction of oxidative deamination of l-amino acids [21].
These proteins have a very wide range of action from anticoagulation
and inhibition of platelet aggregation to anti-viral and anti-bacterial
properties [22,23,24,25]. In obtained gels l-amino acid oxidases appear in two areas (Figure 1 and Figure 2, # 3), and represent 9% of venom proteins. Spots in the upper molecular weight range correspond to the literature data [25]
with weight of approximately 50 kDa. In turn, the spot in the lower
molecular weight region of the gel match only the data from the UniProt
(P0C2D7 (OXLA_VIPBB)) suggesting that this 88-amino acid protein has a
mass of approximately 10 kDa. This observed difference may be due to the
level of protein glycosylation, as in other species, or occurrence of
isoforms of this enzyme [24,26].
The second protein derived from V. berus berus
is a member of cysteine-rich venom protein CRISP (# 6). Proteins from
this group shows wide variety of biological activities. There are many
reports indicating that several venom-derived CRISPs could exhibit
neurotoxicity due to their inhibitory effect on different types of ion
channels [27,28]. Our experiment showed that this is the 4th largest group of proteins in the analyzed venom (6%).
The
only protein that was not found as a result of our experiment was
hyaluronidase. Hyaluronidase causes degradation of hyaluronic acid which
increases the permeability of the tissue at the bite site, and hence
the degree of absorption of the venom. Their action results in local
swelling, blistering and necrosis [3].
Although numerous literature sources indicate that the venom has such
properties, the factor responsible for them has not yet been found.
Interestingly, despite many citations [25,29,30,31] only one work actually states the presence of agents capable of carrying out the depolymerization of hyaluronic acid [32]
Peptidome
analysis showed the presence of 8 peptides in European adder venom, of
which only four could be identified. All of them were identified as
bradykinin -potentiating peptides. Hence, all of them could be inhibitors
of angiotensin-converting enzyme and would enhance the action of
bradykinin, and consequently act as hypotensive agents [33]. Potentially, they could act just like captopril, an oral medication based on the peptide from Bothrops jararaca venom. Interestingly, only one peptide detected in this experiment (1166.5968 m/z) was previously identified in other Vipera species [34], others are of Crotalinae origins (Table 2).
Peptides contained in the venom have great pharmacological potential.
They are poorly immunogenic and have evolutionary conserved tertiary
structure, obtained mostly by disulfide bonds and posttranslational
modifications [35]. The most common of these modifications is pyroglutamate residue at the N-terminus [33,34] observed in two peptides of V. berus berus (Table 2).
It
is necessary to note that the meaningful identification for 2 out of 4
isolated peptides have been obtained only when the posttranslational
modification of deamination NQ was included in the Mascot search
parameters. Unfortunately, it is not possible to determine with our
current experimental setup if such a modification is of a natural origin
or it is an artifact.
Presented results
clearly show that the venom of European adder has mainly hemotoxic
effect, as inferred from a large number of proteins from the family of
metalloproteinases, serine proteases, L-amino acid oxidases or C-type
lectin-like proteins. They exhibit toxic effects on the vascular system,
causing abnormal blood clotting. Furthermore, l-amino
acid oxidases act by causing neuromuscular blockade, and lead to the
destruction of the cells by breaking cell membrane during its
depolarization. In the venom of this snake we also observe a few
proteins responsible for neurotoxicity, these are a cysteine-rich
proteins responsible for the blockade of nerve conduction and
phospholipases A2 possessing both neuro-, myo-, cyto- and
hemotoxic properties. Literature data indicate that the effects of
European adder venom is based mainly on the disorder of homeostasis and
the impairment of blood clotting process, as shown by the presented
results. This work describes for the first time the peptidome of V. berus berus.
Identified peptides potentially have blood pressure lowering properties
and may present a valuable target for further pharmacological
investigations.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}